Claude Gordon Douglas a publié en 1911 une méthode qui est devenue la référence absolue pour mesurer le métabolisme humain pendant la majeure partie du 20ᵉ siècle. Cette méthode porte le nom du sac qu’il a conçu pour recueillir l’air expiré. Elle est encore occasionnellement utilisée aujourd’hui comme référence pour valider les équipements modernes.
Douglas est né à Leicester en février 1882, second fils du chirurgien Claude Douglas. Il a étudié au Wellington College et à la Wyggeston Grammar School, a été admis au New College d’Oxford en 1900, et a obtenu ses diplômes de médecine en 1907. Il a passé le reste de sa vie professionnelle à Oxford, attaché au St John’s College, où il a été fellow de 1907 à 1949. Il a été nommé démonstrateur en métabolisme général en 1919, lecteur en 1937, et professeur en 1942. Il a pris sa retraite en tant que professeur en 1949 à l’âge obligatoire, puis a immédiatement accepté un poste de démonstrateur afin de continuer à enseigner, ce qu’il a fait jusqu’en 1953.
Sa carrière scientifique a été marquée par sa collaboration avec John Scott Haldane, dont Douglas a rejoint le groupe de recherche sur la physiologie respiratoire en 1908. Haldane était la figure dominante de la physiologie respiratoire britannique de l’époque, et la carrière de Douglas s’est construite au sein de son école. Leurs travaux ont porté sur la ventilation, les échanges gazeux, la régulation de la respiration et la physiologie de l’altitude.
En 1910, Douglas a participé à une expédition en haute altitude à Tenerife organisée par le physiologiste allemand Nathan Zuntz, avec Joseph Barcroft de Cambridge et Arnold Durig de Berlin. L’année suivante, il faisait partie de l’expédition anglo-américaine au Pikes Peak, où Haldane, Douglas, Yandell Henderson et Edward Schneider ont passé plusieurs semaines dans la maison au sommet du Colorado pour étudier les effets de la basse pression barométrique sur la respiration. Les résultats, publiés dans les Philosophical Transactions of the Royal Society en 1913 [1], sont devenus des références fondamentales pour la physiologie de l’altitude. Les données de l’expédition ont servi de base à la compréhension de l’acclimatation, de la réponse ventilatoire hypoxique et de la chimie de l’adaptation de la respiration humaine à l’air raréfié.
Le sac
L’article de 1911 qui a valu à Douglas une renommée durable est court — deux pages dans le Journal of Physiology, intitulé « Une méthode pour déterminer l’échange respiratoire total chez l’homme » [2]. Il décrit un appareil suffisamment simple pour être reproduit dans n’importe quel laboratoire : un grand sac en toile, doublé pour limiter la diffusion des gaz, avec un embout buccal, une valve unidirectionnelle et un robinet. Le sujet respire à travers l’embout buccal pendant une période mesurée — quelques minutes, généralement — et tout l’air expiré est recueilli dans le sac. Le volume total est ensuite mesuré, et un échantillon est prélevé et analysé pour déterminer la teneur en oxygène et en dioxyde de carbone. À partir de ces trois chiffres — volume, fraction d’O₂, fraction de CO₂ — on peut calculer la consommation d’oxygène, la production de dioxyde de carbone et le quotient respiratoire, qui décrivent ensemble le métabolisme de l’organisme entier.
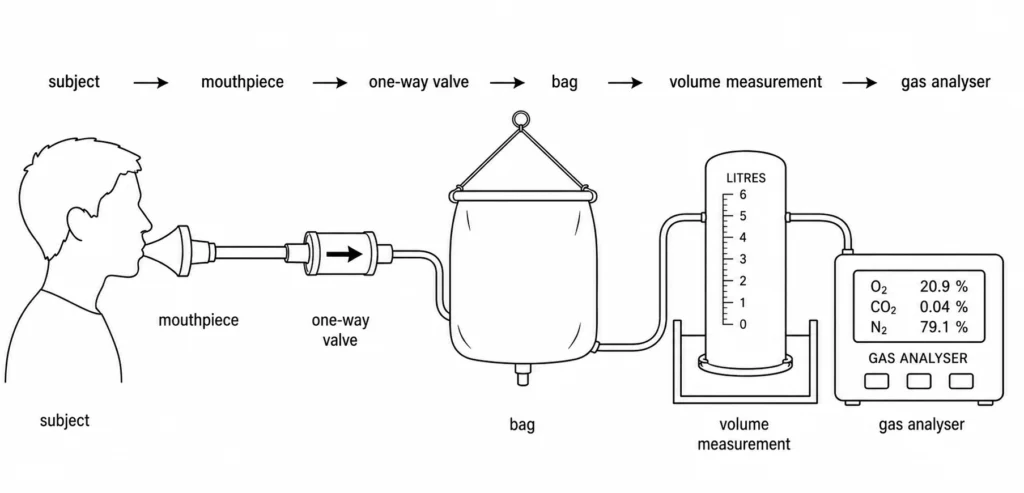
La force de cette méthode résidait dans son caractère direct. Les méthodes antérieures de respirométrie reposaient sur des chambres à circuit fermé (le calorimètre de Lavoisier, le calorimètre de chambre d’Atwater) qui étaient élaborées et coûteuses, ou sur des calculs indirects à partir de mesures incomplètes. Le sac de Douglas plaçait l’appareil sur le sujet et permettait à l’expérience de suivre le sujet — sur un tapis roulant, en montagne, sur le terrain. Il était rudimentaire, mais il fournissait des échantillons complets et sans ambiguïté de gaz expiré.
Ce caractère rudimentaire était important pour une autre raison : la méthode était robuste. Les erreurs sont bien caractérisées et faibles — diffusion des gaz à travers la doublure du sac, contamination de l’espace mort, résistance de la valve — et la plupart ont des corrections connues. Un opérateur méticuleux pouvait mesurer la VO₂ avec une précision de quelques pour cent, ce qui est suffisant pour presque toutes les questions physiologiques.
Cette combinaison — caractère direct, robustesse et faible coût — explique pourquoi le sac de Douglas a survécu aux technologies qui étaient censées le remplacer. Les chariots métaboliques modernes utilisent des analyseurs de gaz à chaque respiration et des débitmètres électroniques ; ils sont plus rapides, plus pratiques et produisent des données en continu. Ils sont aussi imparfaits, et leur étalonnage a historiquement été validé par rapport aux mesures effectuées avec le sac de Douglas. Une revue de 2017 publiée dans l’European Journal of Applied Physiology a passé en revue un siècle d’analyse des erreurs de cette technique et a conclu que, avec les précautions appropriées, elle reste valide en tant que référence standard [3].
Ce qui a survécu
La suite de la carrière de Douglas a été substantielle mais plus discrète que ses années de collaboration. Il a continué à travailler à Oxford sur l’exercice, la respiration et la circulation dans les années 1920 et 1930, a siégé dans des comités gouvernementaux et a formé des générations de physiologistes. L’un de ses derniers étudiants en recherche était Roger Bannister, qui allait courir le premier mile en moins de quatre minutes un an après que Douglas ait finalement pris sa retraite de l’enseignement. Douglas est décédé en 1963, célibataire, à la Radcliffe Infirmary d’Oxford.
Le sac lui-même a eu une étrange postérité. Le modèle original de 1911 — toile, caoutchouc, embout buccal, valve — a été perfectionné mais pas fondamentalement remplacé. Le même appareil de base, avec des matériaux améliorés, est encore vendu aujourd’hui par les fournisseurs de matériel de laboratoire. Des générations de doctorants en physiologie de l’exercice ont effectué leurs premières mesures métaboliques avec un sac de Douglas, selon le principe qu’il faut comprendre la technique en la pratiquant de manière lente avant de faire confiance à la version automatisée.
La méthodologie plus large que Douglas a contribué à établir — la respirométrie en circuit ouvert, la mesure systématique des échanges gazeux pour caractériser le métabolisme — est le fondement de toutes les affirmations modernes concernant la VO₂ max, la dépense énergétique, le métabolisme et la condition aérobie. Les travaux d’A.V. Hill de 1922-23 sur la consommation maximale d’oxygène, dont j’ai parlé séparément, dépendaient de méthodes de collecte de gaz directement issues de celles de Douglas. Il en va de même pour chaque estimation de calories brûlées par les trackers de fitness, chaque test d’effort clinique en cardiopulmonaire, chaque valeur de VO₂ max d’un athlète d’élite. L’instrument a été miniaturisé et numérisé ; le principe n’a pas changé.
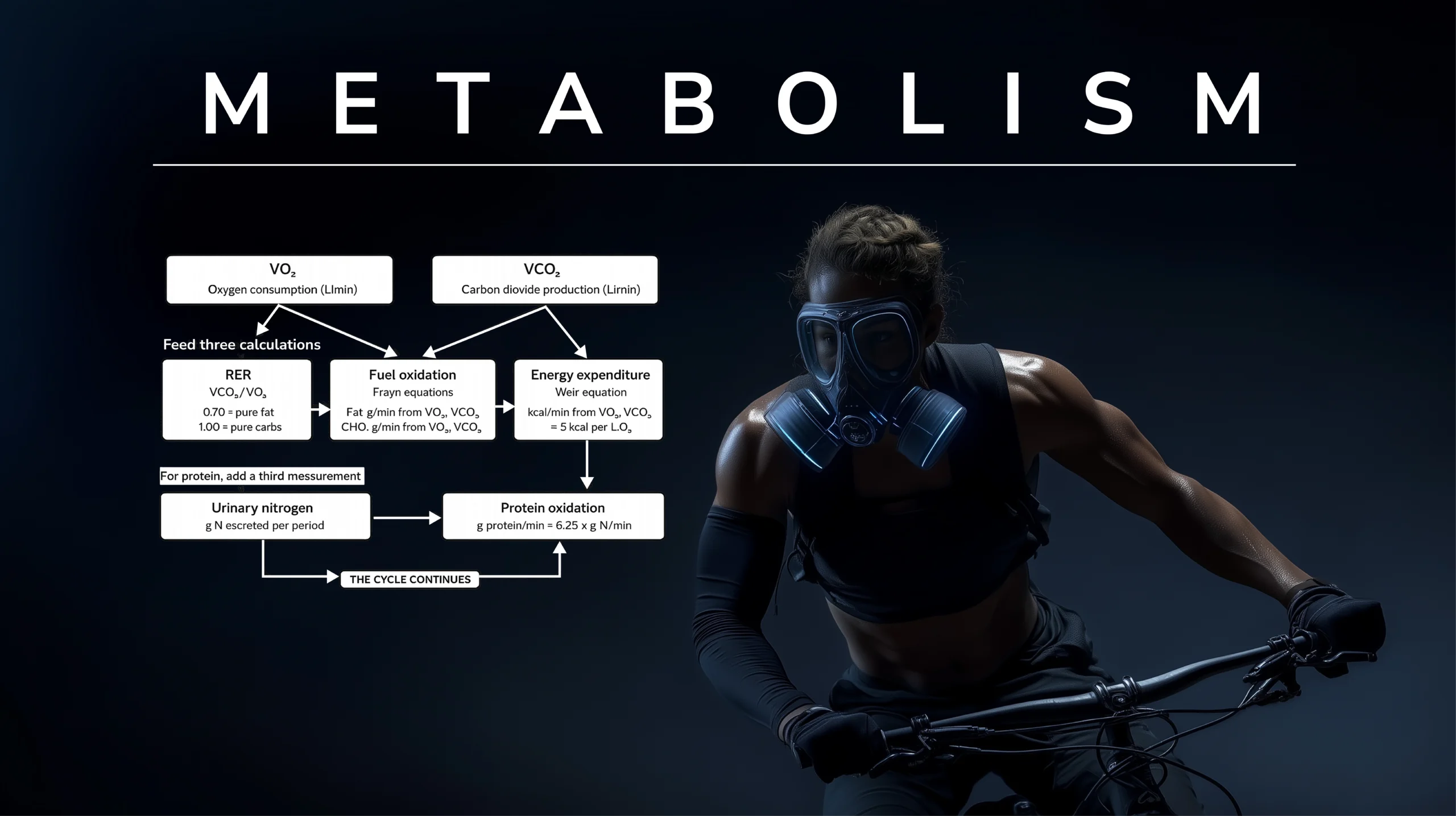
Ce que les deux mêmes mesures — V̇O₂ et V̇CO₂ — permettent de calculer va bien au-delà de ce que suggère l'appareil. Le quotient d'échange respiratoire (RER, le rapport des deux) offre une fenêtre sur le mélange de carburants : un RER d'environ 0,70 indique une oxydation pure des graisses, un RER d'environ 1,00 une oxydation pure des glucides, et les valeurs intermédiaires un mélange dont les proportions peuvent être lues dans des tables standard. Les équations de Frayn (1983) [4] transforment V̇O₂ et V̇CO₂ en grammes par minute de graisses et de glucides oxydés. L'équation de Weir (1949) [5] transforme ces deux mêmes mesures en dépense énergétique totale en kilocalories par minute — le nombre de calories brûlées que tous les trackers de fitness tentent d'estimer depuis.
L'oxydation des protéines est l'exception qui confirme la règle. Son RER (≈0,81) se situe au milieu de la plage graisses-glucides et ne peut pas être distingué par les échanges gazeux seuls. L'équation complète de Weir inclut un terme d'azote urinaire pour la gérer ; la calorimétrie d'exercice aigu fixe généralement ce terme à zéro en supposant que les protéines contribuent de manière négligeable à l'utilisation des carburants au cours d'une courte séance, et le biais résultant est faible. La synthèse des protéines — le tissu en construction plutôt qu'en combustion — n'est pas accessible à partir des échanges gazeux ; elle nécessite des méthodes de traceurs isotopiques stables qui se situent en aval de la spectrométrie de masse plutôt que dans la lignée de Douglas.
Le point essentiel est qu'un seul appareil de 1911 — un sac en toile, un embout buccal, une valve — permet, avec deux raffinements analytiques publiés à des décennies d'intervalle, de rendre compte quantitativement de la quantité de graisses et de glucides qu'un athlète brûle et du nombre de calories qu'il dépense. C'est la portée méthodologique de la contribution de Douglas. Le sac lui-même ne fait pas le travail ; c'est le sac plus Frayn plus Weir qui le fait. Mais rien de tout cela ne serait possible sans le sac.
Dans Afitpilot
Afitpilot se situe une génération méthodologique entière en aval du sac de Douglas — et n'en capture actuellement aucune partie. La plateforme gère la charge de force par le biais d'un 1RM estimé à partir des répétitions par série, du poids et éventuellement du RIR, et gère le stress d'entraînement global par le biais du sRPE de Foster multiplié par la durée. Elle ne dispose d'aucune mesure des échanges gazeux, d'aucune estimation de la VO₂, d'aucune capture de la fréquence cardiaque, d'aucune VRC, d'aucune intégration de wearables et d'aucune représentation de la dépense énergétique en kilocalories ou en kilojoules. Le travail d'endurance entre dans la plateforme sous forme de durée de séance et d'effort perçu, et non sous forme de capacité aérobie ou de cinétique de l'oxygène.
-
RPE de séanceBorg CR10, multiplié par la durée (Foster 2001) → unités arbitraires de charge d'entraînementFoster 2001
-
Répétitions en réservePar série, 0–4. Alimente le calcul du 1RM estimé (Epley) par exercice.Zourdos 2016
-
Disponibilité quotidienneIndice Hooper en cinq points : sommeil, courbatures, fatigue, humeur, stress, chacun noté de 1 à 7.Hooper 1995
-
Charge hebdomadaire en UA + tendance aiguë/chroniqueMoyenne exponentielle pondérée sur des fenêtres de 7 et 28 jours. Purement descriptive — aucun signal de prédiction de blessure.Impellizzeri 2020
-
VO₂ maxAucune mesure directe, aucune estimation, aucune entrée via wearable.Douglas 1911
-
Calories / dépense énergétiqueAucun MET, aucun kJ, aucun champ kcal dans le schéma.—
-
Fréquence cardiaque / VRCAucune fréquence cardiaque au repos, aucune zone, aucune variabilité.—
-
Intégrations de wearablesAucune. Pas d'Apple Health, Garmin, Strava, Whoop, Oura, Polar.—
Cette omission est en partie intentionnelle et en partie simplement liée à l'état actuel de la feuille de route. La partie intentionnelle : la couche d'entrée que la plateforme a construite — sRPE, RIR, disponibilité quotidienne — représente tout ce qu'un athlète peut auto-déclarer sans instrumentation, ce qui maintient le coût d'utilisation du système à zéro matériel. Les entrées instrumentées posent leurs propres problèmes de mesure (variabilité de la fréquence cardiaque de qualité grand public, désaccord entre wearables sur les estimations de VO₂, la vérité terrain bruyante de tout appareil qui n'est pas un chariot métabolique) et des problèmes d'intégration (API des fournisseurs, fiabilité de la synchronisation, l'histoire de la portabilité des données pour les athlètes qui changent d'appareils). Construire correctement la couche wearable n'est pas un petit travail, et la livrer mal introduirait du bruit dans une pile actuellement construite autour d'entrées que l'athlète contrôle.
La partie non intentionnelle : toute la moitié de la physiologie ouverte par la lignée de Douglas — cinétique de l'oxygène, dépense énergétique, plafond aérobie, récupération autonome — se situe en dehors du modèle actuel de la plateforme, et cela ne devrait pas durer éternellement. Une intégration de wearables qui fournirait la fréquence cardiaque, la variabilité de la fréquence cardiaque et une VO₂ max estimée permettrait à la plateforme de représenter l'entraînement d'endurance comme autre chose que « minutes passées à un effort perçu N ». Ce travail figure sur la feuille de route ; il n'est pas encore réalisé aujourd'hui.
J'ai écrit séparément sur A.V. Hill, dont les travaux sur la consommation maximale d'oxygène s'appuyaient directement sur les méthodes de collecte de gaz établies par Douglas.
Références
Références
[1] Douglas, C.G., Haldane, J.S., Henderson, Y., & Schneider, E.C. (1913). Physiological observations made on Pike's Peak, Colorado, with special reference to adaptation to low barometric pressures. Philosophical Transactions of the Royal Society B, 203, 185–318.
[2] Douglas, C.G. (1911). A method for determining the total respiratory exchange in man. Journal of Physiology, 42(Suppl), 17–18.
[3] Shephard, R.J. (2017). Open-circuit respirometry: a brief historical review of the use of Douglas bags and chemical analyzers. European Journal of Applied Physiology, 117(3), 381–387.
[4] Frayn, K.N. (1983). Calculation of substrate oxidation rates in vivo from gaseous exchange. Journal of Applied Physiology, 55(2), 628–634.
[5] Weir, J.B. de V. (1949). New methods for calculating metabolic rate with special reference to protein metabolism. Journal of Physiology, 109(1-2), 1–9.
Laisser un commentaire
Vous devez être connecté pour publier un commentaire.